


La régulation du cycle cellulaire : introduction générale
1- Les différentes phases et les événements du cycle cellulaire
Le cycle cellulaire des cellules des eucaryotes supérieurs comprend quatre phases. Durant deux de ces phases, la phase S et la phase M, les cellules exécutent les deux événements fondamentaux du cycle: la réplication de l’ADN (phase S, pour synthèse) et le partage rigoureusement égal des chromosomes entre les 2 cellules filles (phase M, pour mitose). Les deux autres phases du cycle, G1 et G2, représentent des intervalles entre les deux phases S et M (leur nom vient du mot anglais « gap » qui signifie intervalle). Au cours de la phase G1, la cellule effectue sa croissance, intègre les signaux mitogènes ou anti-mitogènes et se prépare pour effectuer correctement la phase S ; au cours de la phase G2, la cellule se prépare pour la phase M.
Lorsqu’une cellule ne se divise pas, on dit qu’elle est quiescente. On parle alors de phase G0 pour définir cet état non prolifératif. La phase G0 est généralement atteinte après une division mitotique, mais des exceptions existent (dans certains tissus de larves d’insectes ou chez les plantes, certaines cellules entrent en G0 après une phase G2). Les cellules peuvent quitter l’état quiescent pour entrer dans le cycle après une stimulation mitogène.
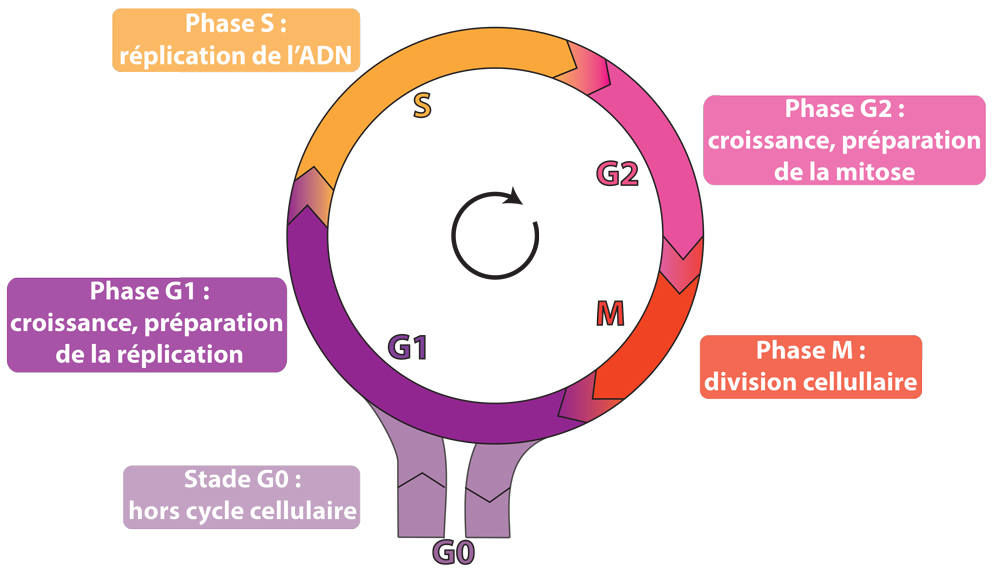
Les quatre phases du cycle cellulaire et la phase G0.
Durant un cycle, les quatre phases se succèdent dans un ordre immuable: G1, S, G2 et M. Les trois premières phases (G1, S, G2) constituent l’interphase, durant laquelle le noyau de la cellule est limité par une enveloppe nucléaire, alors que la mitose (M) est caractérisée par la disparition de cette enveloppe et par l’apparition de la forme condensée de la chromatine, les chromosomes. Ces derniers deviennent alors visibles au microscope photonique parce qu’ils sont compacts.
Après la mitose, les cellules filles peuvent soit entrer en G1 pour effectuer un nouveau cycle de division, soit entrer en G0, phase de quiescence. L’ensemble de ces événements aboutit à la formation de deux cellules filles génétiquement identiques.
2- Comment visualiser les phases du cycle au microscope
La microscopie permet de visualiser certains des événements essentiels du cycle cellulaire, notamment ceux qui sont caractéristiques des phases S et M. En revanche, il est plus difficile de distinguer entre elles les phases Gap.
a. La réplication de l’ADN (c’est à dire des chromosomes) peut être visualisée grâce à la détection d’un analogue de structure de la thymidine (BrdU ou EdU), incorporé dans l’ADN des cellules vivantes au cours d’une exposition courte (pulse).
b. Le centrosome peut être visualisé par immunomarquage à l’aide d’anticorps dirigés contre des protéines centrosomales (par exemple la gamma-tubuline). La duplication du centrosome étant initiée au cours de la phase S, les cellules en G0/G1 possèdent un seul centrosome alors que les cellules en G2/M en possèdent deux.
c. L’enveloppe nucléaire est désorganisée durant la mitose, elle disparait en début de mitose et se ré-assemble en fin de mitose. L’intégrité de l’enveloppe nucléaire peut être suivie par immunomarquage à l’aide d’anticorps dirigés contre des protéines spécifiques de l’enveloppe nucléaire (par exemple le récepteur de la lamine B).
d. Les chromosomes sont condensés pendant la mitose. Ils seront capturés puis alignés avant d’être ségrégés par le fuseau mitotique, un réseau de microtubules dynamiques qui est caractéristique de l’état mitotique. La visualisation des chromosomes (par exemple par coloration de l’ADN) et des microtubules (par marquage de la protéine tubuline) permet de reconnaitre l’état mitotique et les différents stades de la mitose.
e. La cytodiérèse (aussi appelée cytocinèse) est la période ultime de la division cellulaire. Elle est programmée dès la mitose mais s’achève une fois que les cellules filles sont sorties de l’état mitotique, par le clivage du pont cytoplasmique qui les relie encore. Ce clivage est aussi appelé abscission (par analogie avec le clivage du pétiole des feuilles en automne). On peut visualiser le pont cytoplasmique qui relie les cellules en cours de cytodiérèse par marquage de la tubuline, car ce pont est très riche en microtubules.
3- La régulation du cycle cellulaire
Pour assurer, d’une part, l’ordre immuable de la succession des quatre phases du cycle (régulation du cycle), et d’autre part, l’obtention de deux cellules filles rigoureusement identiques (surveillance de l'ADN), la cellule dispose de systèmes de régulation hautement perfectionnés, qui opèrent à différents niveaux.
Dans le premier cas, il existe une machinerie moléculaire qui assure l’exécution de chaque phase mais aussi les transitions entre phases du cycle. Ces moteurs moléculaires sont des protéines enzymatiques, les kinases Cycline-dépendantes, ou Cdk, dont la séquence d’activation puis d’inhibition rythme chaque cycle cellulaire. La succession normale des différentes phases ne peut avoir lieu que si les différentes Cdk intervenant au cours des différentes phases sont présentes et actives aux moments opportuns.
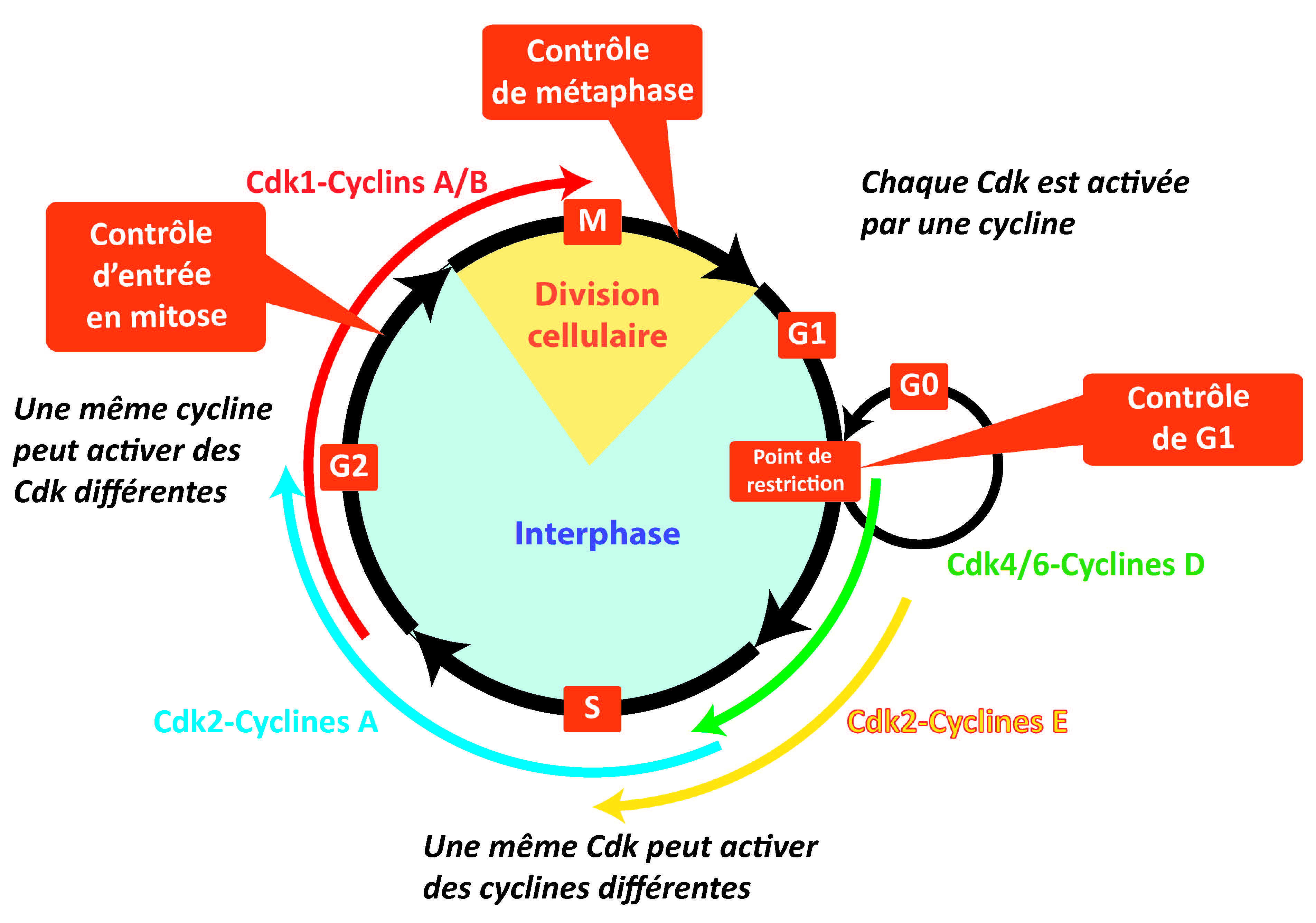
Les moteurs du cycle cellulaire: les complexes Cdk
Dans le second cas, il existe un système de surveillance de la bonne exécution de chaque phase et des transitions entre phases. Un ensemble de mécanismes moléculaires sonde les événements majeurs du cycle (comme un « contrôle qualité ») et peut inhiber les Cdk et donc arrêter le cycle, si l'étape précédente n'est pas terminée, ou si une "réparation" est nécessaire. Ils permettent la surveillance d’aspects fondamentaux comme par exemple l’état des molécules d’ADN avant, pendant et après leur réplication (DDCP = DNA Damage Checkpoint), l’achèvement total de la réplication avant l’entrée en mitose (RCP = Replication Checkpoint) et le bon alignement de tous les chromosomes sur la plaque métaphasique avant la séparation des chromatides-sœurs (MCP = mitotic Checkpoint).
![]()

![]()
